
Bacterial Cell Wall Hydrolysis by Lysozyme PowerPoint PPT Presentation
1 / 16
Title: Bacterial Cell Wall Hydrolysis by Lysozyme
1
Bacterial Cell Wall Hydrolysis by Lysozyme
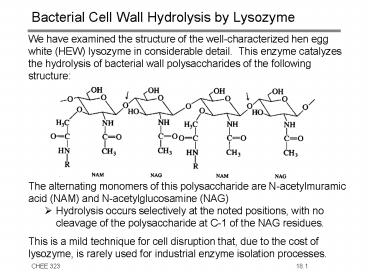
- We have examined the structure of the
well-characterized hen egg white (HEW) lysozyme
in considerable detail. This enzyme catalyzes
the hydrolysis of bacterial wall polysaccharides
of the following structure - The alternating monomers of this polysaccharide
are N-acetylmuramic acid (NAM) and
N-acetylglucosamine (NAG) - Hydrolysis occurs selectively at the noted
positions, with no cleavage of the polysaccharide
at C-1 of the NAG residues. - This is a mild technique for cell disruption
that, due to the cost of lysozyme, is rarely used
for industrial enzyme isolation processes.
2
Characterization of the Active Site
- While the mechanism of every catalytic process is
very difficult to determine, enzyme mediated
reactions are particularly troublesome, as we
have very little knowledge of which functional
groups participate directly in the catalysis. - Kinetic data is useful for design purposes, but
it rarely leads to a reliable reaction mechanism
without supporting information. - Product distribution analysis of model compounds
that approximate the reactive functionality of
the polysaccharide. - Crystallographic analysis of the enzyme and
various enzyme-inhibitor or enzyme-substrate
complexes - Assessment of the influence of select amino acid
substituent modification - Measurement of kinetic isotope effects
- All four of these techniques have been applied to
lysozyme catalysis, and we will examine the first
three to better understand the nature of
enzyme-substrate interactions.
3
Lysozyme Activity Studies - Model Compounds
- The cleavage patterns for acetylglucosamine
oligomers - are not consistent with a random attack of the
enzyme. - Cleavage occurs at an appreciable rate only for
hexamers or higher oligomers and occurs between
mers 4 and 5. - Indicates that a hexasaccharide is incorporated
by the catalytic site of the enzyme, and a unique
mode of substrate activation must establish a
preferred hydrolysis pathway.
4
Crystallographic Studies of Lysozyme-Inhibitor
Complexes
- Once the crystal structure of an enzyme has been
determined the structures of isomorphous crystals
that contain additional molecules may be
determined without difficulty. - This method has been used to explore the
interactions between lysozyme and a wide range of
substrate-related inhibitor molecules.
Perspective drawing of the main chain
conformation of lysozyme Elevation from
active-site side of the molecule. Only a
positions of a-carbon atoms are shown.
5
Crystallographic Studies of Lysozyme-Inhibitor
Complexes
- The trisaccharide of NAG forms a relatively
stable complex with lysozyme that has been
characterized by crystallography. The position
of the three NAG groups is illustrated at the
top-right of the structure. - The other three groups have been
- positioned by molecular
- modeling, as they cannot be
- isolated.
- N-acetylglucosamine (NAG) alone
- binds to the enzyme with H-bonds between the NH
and carbonyl - oxygens of its acetamido side chain and the main
peptide chain and CO and NH groups of residues
107 and 59.
6
Active Site Determination Chemical Modification
- If the chemical modification of a particular
amino acid side chain results in enzyme
deactivation, then the residue in question is
located at the active site, provided that the
modification can be prevented by the presence of
excess substrate or inhibitor. The following is
a summary of specific modifications used to
determine the activity of individual residues - A. Amino (Lysyl e-amino, 1,13, 33, 96, 97,116)
- All 6 lysine residues are on the surface of the
enzyme, with Lys 33 situated in the very bottom
of the cleft. - Little effect of modifications of this type on
lytic activity is observed, suggesting that these
amino acid residues do not participate directly
in catalysis.
7
Active Site Determination Chemical Modification
- B. Arginine (5, 14, 21, 45, 61, 68, 73, 112, 114,
125, 128) - All but one Arg is located on the surface of the
enzyme, but Arg 114 is believed to form two
hydrogen bonds with the saccharide. - Modification of 7 of the 11 Arg residues had
little influence on the activity of lysozyme on
NAG4. - C. Glutamic Acid 7, 35 Aspartic Acid 18, 48,
52, 66, 87, 101, 111 - Modification studies of carboxyl groups have
provided unequivocal evidence for the involvement
of these residues in catalysis. - Exhaustive esterification of lysozyme with acid
alcohol results in a loss of enzyme activity
8
Active Site Determination Chemical Modification
- C. Carboxyl Groups Continued
- Two amino acid residues in particular (Asp 52 and
Glu 35) have been implicated in several
modification studies. - Modification of Asp 52 was inhibited by the
presence of substrate - Selective oxidation of Glu 35 with iodine
denatures the enzyme. - D. Cysteine (6-127, 30-115, 64-80, 76-94)
- Reduction of disulfide crosslinks denatures the
enzyme, although 6-127 can be opened without
deactivation. This is reversible, as air
oxidation regenerates enzymatic activity with
high yield. - E. Histidine (15)
- Alkylation of the single histidine has been shown
to have little influence on lytic activity.
9
Active Site Determination Chemical Modification
- F. Methionine (12,105)
- Cyanogen bromide in 70 formic acid cleaves both
methionyl peptide bonds without modification of
other amino acid sequences. - This reduces activity to 10 of the native
enzyme, despite the fact that both residues are
buried within the enzyme structure and
participate through non-polar contacts with other
residues. - G. Tryptophan (28, 62, 63, 108, 111, 123)
- Three of six tryptophans are believed to be
positioned in the active site. Oxidation with
N-bromosuccinimide inactivates the enzyme. - Selective oxidation of Trp 108
- with iodine is blocked by
- substrate. Trp is also in close
- proximity to Glu 35.
10
Binding of Lysozyme to Hexa-N-Acetylglucosamine
- Schematic illustration of the active site in the
cleft region of lysozyme. A through F represent
the glycosyl moieties of a - hexa-saccharide. Some of the amino
- acids in the cleft region near
- these subsites of the
- active site
- are shown.
11
Model of the HEW Lysozyme Site
- Schematic diagram showing the specificity of
lysozyme for hexa-saccharide substrates. - Six subsites A-F on the enzyme bind the sugar
residues. Alternate sites interact with the
acetamido side chains (a), and these sites are
unable to accommodate MurNAe residues with their
lactyl side chains (P). - Site D cannot bind a sugar residue without
distortion, and the glycosidic linkage that is
cleaved binds between sites D and E as shown by
the arrow.
12
Proposed Catalytic Mechanism
- 1. The saccharide binds in the enzyme cleft with
residue D distorted to a conformation resembling
the half-chair. - 2. Bond rearrangement to yield a carbenium ion
proceeds at a rate enhanced through several
contributions - A. Glu 35 acts as a general acid catalyst,
donating H to the glycosidic oxygen. - B. Asp 52 bears a negative charge that favors
formation of the carbenium ion. - C. The ring conformation is close to that
required in the transition state. - D. The nonpolar nature of the cleft possibly
enhances reaction rate. - 3. The enzyme-bound carbonium ion is stabilized
by neighboring charges of Asp 52 and Glu 35, the
latter having deprotonated in bond rearrangement.
- 4. The aglycone diffuses away, and reaction with
water or another acceptor completes the process.
13
pH Dependence of Enzyme Activity
- Since the characteristics of ionizable side
chains of amino acids depend on pH, enzyme
activity varies with pH shifts. - At extremes of pH, the tertiary structure of the
protein may be disrupted and the enzyme
denatured. - Even at moderate pH values where tertiary
structure is unaffected, enzyme activity may
depend on the degree of ionization of certain
amino acid side chains - pH can therefore affect enzyme
- conformation, substrate binding
- and the ability of active side
- groups to participate in catalysis,
- as shown here for three
- representative enzymes.
14
Survey of Ionizable Enzyme Groups
- The ionizable groups which contribute to the
acid-base properties of proteins, shown with
their approximate pKa values. These can vary by
several pH units depending on their environment
in the protein.
15
pH Dependence of Enzyme Activity
- Recall our discussion of lysozyme, where Asp52
was believed to exist in its conjugate base
(RCOO-) form, while Glu35 is thought to be active
in its acidic state (RCOOH). - pH will dictate the degree of protonation of
these residues, creating an optimum that is
dependent on their pKas. - Graphs of Vmax against pH, at constant E, where
catalytic activity depends on the simultaneous
presence of EY- and EZH - (a) where pKy and pKz are more than 2 units apart
- (b) where pKy and pKz are less than 2 units apart
16
pH Dependence of Enzyme Activity
- Consider a reaction that requires the conjugate
acid/base pair HA/A- to be in its basic form, and
the conjugate acid/base pair HB/B- to be in its
acidic state. The system speciation will change
with pH, as follows.
Recommended
