
Conditional Specification PowerPoint PPT Presentation
1 / 38
Title: Conditional Specification
1
Conditional Specification
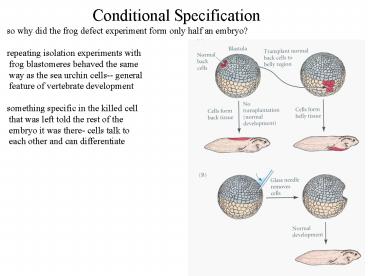
so why did the frog defect experiment form only
half an embryo? repeating isolation experiments
with frog blastomeres behaved the same way as
the sea urchin cells-- general feature of
vertebrate development something specific in the
killed cell that was left told the rest of the
embryo it was there- cells talk to each other
and can differentiate
2
Autonomous Specification
morphogenic determinants something in the cell
which determines type ie. polar granules in C
elegans determine germ cells of the embryo in
tunicates, cells can be taken out of the embryo
and will form the same type of cell as they
normally would
EMS
P2
ABa
ABp
presumptive trochoblasts remember what they
should become, even in isolation
3
Morphogenic Gradients
morphogen soluble instructive molecule secreted
at a distance from the target when a
substance is secreted, it diffuses so that there
is a high local concentration and decreasing
concentrations at greater distances cells in
different parts of this concentration gradient
can receive different signals and become
different cell types Ephrins are signaling
molecules that regulate the formation of
connections in the brain
4
Morphogenic Gradients
same holds true for extracellular gradients as
shown in flatworms cutting off the heads or the
tails of flatworms both halves regenerate
the missing half, not the same half leave most
of the middle, both front and back
regenerate with only a small part of the middle
the regeneration fails
5
Morphogenic Gradients
for a substance to be considered a morphogen,
cells must respond directly to that substance
based on its concentration activin a protein
from frogs that affects different cells based on
activin increases gene expression related to
the formation of mesoderm has several related
TGF-b signaling molecules like nodal (below)
mouse nodal in situ
6
Morphogenic Gradients
concentration of activin causes blastomeres to
adopt different fates
7
Morphogenic Gradients
morphogenetic field group of cells where
position within certain boundaries determines
the cell fate ie. imaginal disks in flies stay
imaginal disks no matter where they are
transplanted in an embryo-- they can become leg
or wing or antenna,etc but individual cells
are not committed to a particular cell
type cells within a morphogenetic field interact
among themselves to determine what type of
cell within a field they will become
8
Cell and Tissue Morphology
2 major types of cell arrangements in the
embryo 1) epithelial cells-- tightly connected
sheets or tubes of cells 2) mesenchymal cells--
independent, unconnected cells
epithelial cells mesenchymal cells
9
Cell and Tissue Morphology
within these 2 types of arrangements there are
several ways cells organize a)
direction/orientation of cell divisions b) cell
shape changes c) cell movement/migration d)
cell growth e) cell death-- not every cell born
lives f) change membrane or secreted products
10
Differential Cell Affinity
Cells interact with each other almost exclusively
at the cell surface proteins on the cell
surface recognize other cells, receive signals
such as morphogens, and send out their own
signals selective affinity preferences of
particular cell types/ germ layers to stick to
like cells rather than unlike
11
Differential Cell Affinity
additionally, early aggregates assume the same
positions in vitro as they would in the
embryo-- epithelial cells on the outside,
endoderm inside, mesoderm in
between histotypic aggregation organization of
complex tissue from single cells
Selective affinity changes during development-
cells move and change over time- later cells do
not behave the same as early ones
12
Differential Cell Affinity
differential adhesion hypothesis cells stick
together in order to make the most stable
overall structure (ie. cells alway move toward
lower energy) strongest interactions would most
compact the next strongest, then the next,
etc. like cells that stick together better
than they stick to other cell types will
effectively group together because they stick
when they encounter others pigment epithelium
12.6 dynes/cm neural retina 1.6 dynes/cm sort
very clearly, as in figure
13
Cell Adhesion Molecules
Cadherins large family of calcium dependent
adhesion molecules bind homophilically to each
other-- N with N, E with E, etc transmembrane
proteins that all bind to the same intracellular
proteins catenins complex of 3 proteins which
anchor the cadherins to actin at least one of
these proteins (b catenin) can also act as a
signal a catenin binds directly to actin
cytoskeleton E cadherin initially on all
embryo cells, becomes restricted to epithelial P
cadherin primarily on tropoblasts (placenta) and
uterine wall N cadherin initially on mesodermal
cells, primarily expressed on neurons C
cadherin expressed on blastomere cells and
critical for gastrulation
14
Cell Adhesion Molecules
homophilic binding occurs at the N terminus-- C
terminus is in the cytoplasm calcium ions hold
the domains as a rigid rod-- required for
good adhesion mediates a lot of early adhesion
during development expressing more P cadherin on
the surface makes stronger interactions and
causes those cells to be on the inside of a
group of cells
15
Cell Adhesion Molecules
tropoblasts early cells of embryos that will
form part of the placenta express both P and E
cadherins P cadherins allow the embryo to bind
to the uterine wall in mammals-- required for
the embryo to survive E cadherins allow the
tropoblasts to bind to the rest of the embryo,
anchoring the cells to the uterus
16
Cell Adhesion Molecules
other families of adhesion molecules play vital
roles in development integrins a/b dimers that
recognize distinct amino acid sequences on
exposed regions of cell surface proteins-- RGD is
the most common link to specific kinases in
the cytoplasm kinase protein that
phosphorylates other (usually specific)
proteins immunoglobulin (Ig) superfamily
largest class of adhesion molecules bind both
homophilically (like molecules) and
heterophilically (unlike) contains multiple (1
to 20) repeats of the protein domain in
antibodies links to a variety of cytoplasmic
proteins, including kinases and actin vital
roles best understood in the immune and nervous
systems lectins class of proteins that bind to
specific carbohydrate (sugars) groups bind
various glycoproteins (ie. gp80 in
dictyostelium) are vital for immune cell
function and recognition, also found elsewhere
17
Cell-Cell Communication
induction short range interaction that causes
changes in the adjacent cell-- essential for
forming the correct pattern of cells in organs
2 components the signal, and the target tissue
with the correct receptor competence ability to
respond to an inductive signal head ectoderm
responds to the optic vessicle to induce the
lens abdominal ectoderm does NOT respond, and
no lens tissue formed competence and inductive
ability are both acquired and
maintained Pax6 is required in the ectoderm
knockout mouse optic vesicle forms eye tissue
normally knockout ectoderm cells cannot
respond to normal optic vesicle
18
Cell-Cell Communication
Pax6 by itself is not suffcient to make an eye--
requires other inducers two inducing factors
BMP-4 (bone morphogenetic protein) and FGF-8
(fibroblast growth factor) many induction events
are symmetrical-- the tissue induced to form
signals back to the original source of the
signal reciprocal induction condition where the
induced tissue signals back to the original
inducing cells the same inductive signal can
operate independently on different cell
types-- ie. it has a receptor for an inducer, but
responds differently to that inducer based
upon its differential gene expression
19
Cell-Cell Communication
instructive signal a signal from the inducing
cell is required to cause a particular change
in gene expression to occur several
characteristics of inductive signals in the
presence of the inducing tissue, the responsive
tissue reacts in the absence of the inducing
tissue, the responsive tissue fails to react
in the absence of one inducer but in the presence
of another, the responsive tissue fails to
react correctly permissive signal the tissue
contains what it needs to respond by itself,
but it requires the appropriate environment to
express those traits ie. in the absence of
additional signals, it will develop in a single
way
20
Eye Development
optic vesicle induces ectoderm to express Otx2,
Pax6, Sox3 sequentially neural retina induced by
the ectoderm, with the rest forming pigment
cells lens tissue invaginates (folds into) optic
cup, forms capsule and vesicle lens tissue also
induces the ectoderm to form the transparent
cornea
21
Epithelial- Mesenchymal Interactions
epithelial cells any cells formed into sheets or
tubes-- any germ layer mesenchymal cells loosely
packed, unconnected cells all organs contain
both epithelial and a mesenchymal component
usually has to have something containing the
cells keeping it together skin is made up of
epidermis (ectoderm) and dermis
(mesoderm) epidermis induces condensations in
the dermis, changing it uses secreted proteins
to induce dermis the condensations then induce
the epidermis to form regional
structures epidermis can make any feather, but
the signal from the dermis tells it what to make
22
Developmental mechanisms (not outcomes) are
conserved
cells respond to a particular signal, but the
response will depend upon what type of cell
will do the responding ie. frogs and newts form
very different mouth organs from the same signal
transplant frog ectoderm to newt and newt
ectoderm to frog ectoderm forms a mouth, but
forms the mouth of the donor tissue
23
Inducing Factors
two different ways molecules can act as
inducers juxtacrine factors requires a signal
attached to one cell to interact with a
receptor on another cell-- direct cell-cell
contact filters placed between 2 cells can
block the induction paracrine factors a
diffusable signal moves a short distance and
interacts with a receptor on another cell--
local interaction, but no contact classical
secreted developmental inducer filters placed
between 2 cells cannot block the
induction endocrine factors hormones that
travel through the blood to exert their
effects-- very distant interaction autocrine
factors unusual cells that secrete the factor
also responds to it typically cause a rapid,
extensive result-- ie. volvox sexual induction
24
Paracrine Factors
include 4 major families of proteins-- fibroblast
growth factors, Wnt family, hedgehog proteins,
and transforming growth factor superfamily also
includes several vital but less conserved
proteins epidermal growth factor (drosophila
eye development, worm vulva) stem cell factor
neurotropins (growth/differentiation/survival
factors for neurons) erythropoetin--
stimulates red blood cell production-
chemotherapy cytokines-- stimulates blood cell
differentiation/growth interleukins- usually T
cell factors, possibly increases antiviral
response diffusable factors play some role in
almost all aspects of development
25
Fibroblast Growth Factors
composed of about 2 dozen different, structurally
related proteins different factors affect
different events RNA splicing and alternative
start codons can generate hundreds of
isoforms in a tissue specific fashion FGF2 (aka
basic FGF) is important in angiogenesis and
neural stem cell proliferation FGF8 is expressed
in the optic vesicle which contacts the
ectoderm becomes concentrated in neural
retina FGF8 beads induce head ectoderm to form
ectopic lenses
26
Receptor Tyrosine Kinase Pathway
27
Hedgehog Signal Transduction Pathway
28
Wnt Signal Transduction Pathway
29
TGF-b Paracrine Factor Superfamily
30
TGF-b Signal Transduction
31
Jak-STAT Signal Transduction
32
Pathways of Apoptosis
apoptosis programmed cell death as part of
normal development only about 1/3 of neurons
that were born in embryos survive we recycle
about our own body weight in cells per
year different signals regulate different
pathways of apoptosis-- sometimes signals for
differentiation in one tissue (BMP-4) cause
apoptosis in a different tissue (ie. BMP-4
causes apoptosis in tooth development) erythropoe
itin prevents destruction of red blood cell
precursors in liver in worms, ced-3 and ced-4
are essential for apoptosis (turned off by
ced-9) ced-4 is a protease that activates
ced-3 ced-3 kills the cell excess ced-9
causes excess cell survival deletion excessive
cell death these ced genes are the central
components in all apoptotic pathways
orthologous genes control cell death in flies,
mice, humans, etc
33
Pathways of Apoptosis
Bcl-2 family of genes are the homologs of ced-9
bind to and inhibit ced-4 protease homologs so
it cannot activate death Bcl-2 can directly
substitute for the C elegans gene (very
conserved) Apaf1 is the mammalian homolog of
ced-4 can be activated in several ways
paracrine/endocrine signal, loss of
adhesion, etc works through a cytochrome-c
dependent mechanism to activate ced-3 knockout
mutant mice have massive cell overgrowth and
webbed digits caspases are the homologs of the
ced-3 genes in mammals caspases 3 and 9 are
strong proteases that digest the cell from
inside knockout mutants in either gene causes
massive cell overgrowth and lethality just
near birth different caspases can be activated
by alternative mechanisms depending upon what
tissue they are in- function similarly to C
elegans mechanism
34
Pathways of Apoptosis
35
Juxtacrine Signaling
juxtacrine signaling cells respond to signals
from directly neighboring cells-- no diffusion
is allowed-- and is blocked by filters 3 types
of juxtacrine signaling 1) Membrane attached
signal on one cell binds to a membrane attached
receptor on another cell 2) Membrane
attached receptor binds to the extracellular
matrix made by another cell
extracellular matrix insoluble secreted material
that fills in the gaps between cells--
usually fibrous in nature ie. collagen, laminin,
etc 3) Direct cytoplasm to cytoplasm signaling
through gap junctions juxtacrine and paracrine
signaling systems are NOT exclusive-- often
have both types of signaling going on in a single
tissue
36
Notch Pathway- Surface to Surface Signaling
Notch cell surface protein with an extracellular
and an intracellular part when Notch binds its
ligand extracellularly, a protease in the
cytoplasm cleaves notch, and the cut portion
moves to the nucleus (similar to Ci) same
protease that cleaves Notch cleaves presenilin-1
in Alzheimer's in the nucleus, cleaved notch
binds to inactive transcription factors of the
CSL family, activating them and turning on new
genes Notch binds to any of several ligands
Delta, Jagged, or Serrate
37
Extracellular Matrix and Signaling
38
Gap Junctions and Signaling
gap junction channel connecting two adjacent
cells by means of a protein bridge-- each cell
has a hexamer of connexin proteins binding the
other passes small molecules and ions
freely different connexin proteins form gap
junctions with different and often overlapping
roles in development many (but not all)
functions can be taken over by other
connexins very common in blastulas and
epithelial sheets cleavage communication via
gap junctions is vital in development
Recommended
