
Pr PowerPoint PPT Presentation
1 / 1
Title: Pr
1
Role of the spleen in leukemogenesis induced by
bovine leukemia virus
Florins A 1,
Debacq C 1, Gillet N 1, Jean G 1, Thewis A 1,
Schwartz-Cornil I 2, Bonneau M 2, Hay J 3,
Asquith B 4, Burny A 1, Kettmann R 1 and Willems
L 1 1Molecular and Cellular Biology, FUSAGx,
Gembloux, Belgium 2INRA, Jouy-en-Josas, France
3Immunology, University of Toronto, Canada 4
Dept of Immunology, Imperial College of London,
United Kingdom
We characterized the role of the spleen during
infection of sheep by bovine leukemia virus
(BLV). Experiments based on CFSE labeling showed
that B lymphocytes from infected animals
disappear faster from the blood compartment
compared to the controls. Mathematical modeling
of these data showed that this process was
associated with increased cell death rates
occurring in infected sheep. This difference in
dynamics seemed mainly to be due to the B-CD11b
subpopulation. These cells are indeed excluded
from the lymphatic compartment and migrate
preferentially through the spleen. To study the
involvement of the spleen in the kinetics of
BLV-infected B lymphocytes, we analyzed the
phenotype and proportion of different cell
populations as well as the proviral loads in the
splenic artery and vein of 2 infected sheep. No
difference was observed amongst all measured
parameters. We next performed a kinetic analysis
of the B cells based on the use of intravenous
CFSE injection within 4 splenectomized sheep, two
of them being infected with BLV. Interestingly,
in the absence of spleen, the cell death rates
were similar in BLV-infected and control sheep.
This report thus enlightens a key role exerted by
the spleen in the B cell turnover of BLV-infected
animals.
A.
B.
of B-CD11b in the B lymphocyte population
of B lymphocytes in
the PBMC population
of B lymphocytes in
the PBMC population
4533
4534
4535
2672
4533
4534
4535
2672
4533
4534
4535
2672
4533
4534
4535
2672
Control sheep
Infected sheep
Control sheep
Infected sheep
Control sheep
Infected sheep
Control sheep
Infected sheep
C.
D.
Average numbers of viral
Average numbers of viral
copies per cell
copies per cell
of cells expressing p24
of cells expressing p24
in the PBMC population
in the PBMC population
4535
2672
4535
2672
4535
2672
4535
2672
Jugular vein
Splenic artery
Splenic vein
Jugular vein
Splenic artery
Splenic vein
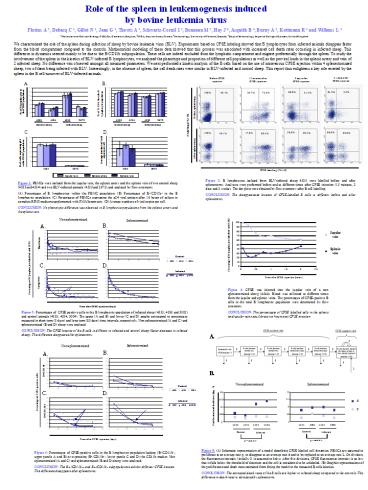
Figure 1 PBMCs were isolated from the jugular
vein, the splenic artery and the splenic vein of
two control sheep (4533 and 4534) and two
BLV-infected animals (4535 and 2672) and
analyzed by flow cytometry. (A) Percentages of B
lymphocytes within the PBMC population. (B)
Percentages of B/CD11b in the B lymphocyte
population. (C) Percentages of PBMCs expressing
the p24 viral antigen after 16 hours of culture
in complete RPMI medium supplemented with
PMA/Ionomycin. (D) Average numbers of viral
copies per cell. CONCLUSION No phenotypic
difference was observed in B lymphocyte
populations from the splenic artery and the
splenic vein.
Non splenectomized
Splenectomized
A.
B.
100
90
Jugular
80
Jugular
vein
70
vein
60
Short term
Short term
Percentage of B lymphocytes labelled with CFSE
50
Splenic
Splenic
40
vein
vein
30
20
D.
C.
10
0
Percentage of B lymphocytes labelled with CFSE
0
1
2
Time after CFSE injection (hours)
Long term
Long term
Figure 4 CFSE was injected into the jugular vein
of a non splenectomized sheep (4544). Blood was
collected at different times from the jugular and
splenic veins. The percentages of CFSE-positive B
cells in the total B lymphocyte population were
determined by flow cytometry. CONCLUSION The
percentages of CFSE labelled cells in the splenic
and jugular veins equilibrate two hours post CFSE
injection
Time after CFSE injection (days)
Time after CFSE injection (days)
Figure 3 Percentages of CFSE positive cells in
the B lymphocyte population of infected sheep
(4535, 4536 and 3002) and control animals (4533,
4534, 3004). The upper (A and B) and lower (C and
D) graphs correspond to percentages measured in
short term (5 days) and long term (83 days) time
intervals, respectively. Non splenectomized (A
and C) and splenectomized (B and D) sheep were
analyzed. CONCLUSION The CFSE kinetics of the B
cells is different in infected and control sheep
(faster decrease in infected sheep). This
difference diappears after splenectomy.
A.
Splenectomized
Non splenectomized
B.
A.
B.
B/CD11b-
Splenectomized
Non splenectomized
0.2
D.
C.
Percentage of CFSE-positive cells
0.15
d
Proliferation and death rates (day 1)
0.1
p
B/CD11b
0.05
0
4533
4535
4534
4536
4534
3004
3002
4535
Control
Control
Infected
Infected
p and d
p and d ?
Time after CFSE injection (days)
Figure 6 (A) Schematic representation of a model
describing CFSE labeled cell dynamics. PBMCs are
assumed to proliferate at an average rate p, to
disappear at an average rate d and to be replaced
at an average rate L. On division, the
fluorescence intensity (initially J) is assumed
to halve. After five divisions, CFSE fluorescence
intensity is so low that it falls below the
threshold of detection and the cell is considered
to be unlabeled. (B) Graphic representations of
the proliferation and death rates estimated from
fitting the model to the measured B cells
kinetics. CONCLUSION The estimated death rates
of the B cells are higher in infected sheep
compared to the controls. This difference in
death rates is abrogated by splenectomy.
Figure 5 Percentages of CFSE-positive cells in
the B lymphocyte population lacking (B/CD11b-
upper panels A and B) or expressing (B/CD11b
lower panels C and D) the CD11b marker. Non
splenectomized (A and C) and splenectomized (B
and D) sheep were analyzed. CONCLUSION The
B/CD11b and B/CD11b- subpopulations exhibit
different CFSE kinetics. This difference
disappears after splenectomy.
Recommended
