
IV. Translation - PowerPoint PPT Presentation
1 / 46
Title: IV. Translation
1
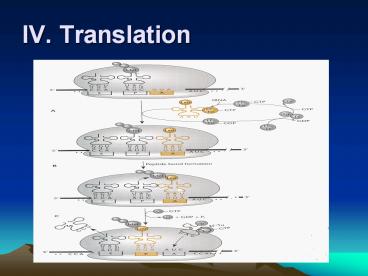
IV. Translation
2
IV. Translation
- (3) Translation termination
- i. When a ribosome comes to a nonsense
codon (or stop - codon, usually one of UAA, UAG and
UGA), translation - stops and polypeptide is released from
ribosome. - ii. Stop codons do not encode an amino
acid, so they have no - corresponding tRNA.
- iii. Termination requires release factors
(RF1 and RF2) which - recognize nonsense codon and promote
the release of the - polypeptide form the tRNA and the
ribosome from the - mRNA. (Fig. 2.35)
3
Termination of translation at a nonsense codon
4
IV. Translation
- 5. Polycistronic mRNA
- ? In bacteria and archaea, the same mRNA can
encode more - than one polypeptide. Such mRNAs,
called polycistronic - mRNAs, must have more than one TIR to
allow simultaneous - translation of more than one sequence
of the mRNA. - (1) Even if the two coding regions overlap,
the two - polypeptides on an mRNA can be
translated independently - by different ribosomes.
- (2) Translational coupling The translation
of upstream gene is - required for the translation of the
gene immediately - downstream. The secondary structure
of the RNA blocks - translation of the second polypeptide
unless it is disrupted - by a ribosome translating the first
coding sequence.
5
Structure of a polycistronic mRNA
- Even if the two coding regions overlap, the two
polypeptides on - an mRNA can be translated independently by
different ribosomes.
6
IV. Translation
- (3) Polar effect on gene expression - Some
mutations that affect - the expression of a gene in a
polycistronic mRNA can have - secondary effects on the expression of
downstream gene. - i. The insertion of an transcription
terminator prevents the - transcription of downstream gene.
- ii. The mutation changing a codon to a
nonsense codon will - dissociate the ribosome from mRNA,
then the translation - of downstream gene that is
translationally coupled to the - upstream gene will not translated.
- (4) ?dependent polarity (as shown in
Fig.2.38) - A. Normally the rut site is masked by
ribosome translating the - mRNA of gene Y.
- B. If translation is blocked in gene Y
by a mutation that changes - the codon CAG to UAG, the ?
factor can cause transcription - termination before the RNA
polymerase reach gene Z. - C. Only the fragment of gene Y protein
and mRNA are produced - and even gene Z is not even
transcribed into mRNA.
7
Model for translational coupling in polycistronic
mRNA
8
Polarity in transcription of a polycistronic mRNA
transcribed from PYZ.
9
V. Regulation of gene expression
10
1. Transcriptional regulation
- (1) Genes whose products regulate the
expression of other - genes are called regulatory genes.
Their products can be - either activator or repressor.
- (2) The set of genes regulated by the same
regulatory gene - product is called a regulon. If a gene
product regulates its - own expression, it is said to be
autoregulated. - (3) Bacterial genes are often arranged in an
operon which - consists of a promoter region, an
operator region and - several structure genes. The mRNA of
bacteria are made on - a number of genes whose products
perform related - functions. This kind of mRNA is called
polycistronic mRNA.
11
Transcriptional regulation
- (4) There are two general types of
transcriptional regulation - i. In negative regulation, a repressor
binds to an operator and - turns the operon off by preventing RNA
polymerase from - using or access the promoter. An
operator sequence can - be close to (up- or downstream), or
even overlapping the - promoter.
- ii. In positive regulation, an activator
binds to the upstream of - the promoter at an upstream activator
site (UAS), where it - can help RNA polymerase bind to the
promoter or help - open the promoter after the RNA
polymerase binds.
12
Two general types of transcriptional
regulation
13
Lac operon
- A. Bacteria respond to rapidly changing
environments - B. Examples
- a. Lac operon(?????)of E. coli(????)
- 1. promoter sequence(?????) RNA polymerase
- (RNA???)
- 2. operator sequence(?????) repressor
- protein
- 3. structural genes (????) Z (beta-
- galactosidase ??????), Y (permease
???) - and A (transferase ???)
- b. regulator gene(????) repressor protein
- (?????)
14
The regulation of gene expression of Lac operon
Operon
Operator
Regulatory gene
Promoter
Lactose-utilization genes
DNA
mRNA
Protei repressr
RNA polymerasecannot attach topromoter
Activerepressor
OPERON TURNED OFF (lactose absent)
DNA
RNA polymerasebound to promoter
mRNA
Protein Repressor
Inactiverepressor
Enzymes for lactose utilization
Lactose
OPERON TURNED ON (lactose inactivates repressor)
15
The ß- galactosidase reaction
16
The lac control region
17
Diauxic growth curve of E. coli grown with a
mixture of glucose and lactose
18
The interaction of promoters and CAP proteins in
Lac operon
- A. CAP proteins are involved in positive
regulation - a. positive regulation activator(???)
- b. CAP catabolite activator
proteins(????????) - c. CAP binding site
- d. cAMP cyclic adenosine monophosphate(??????
?) - e. CAP/cAMP complex increasing the
efficiency the - ability of RNA polymerase binds to
promoter. - B. Catabolite repression(?????) enabling E.
- coli to use glucose (???) preferentially for
- energy even in the presence of lactose or
other - complex sugar.
- a. decreasing the level of cAMP
- b. permease - nonfunctional
19
Positive Control of lac Operon
- Positive control of lac operon by a substance
sensing lack of glucose that responds by
activating lac promoter - The concentration of nucleotide, cyclic-AMP,
rises as the concentration of glucose drops
20
The phosphoenolpyruvate (PEP)-dependent sugar
phosphotransferase system (PTS)
- Both HPr and IIA are the components of the PTS,
which is responsible for transporting certain
sugars, including glucose.
21
Catabolite repression of the lac operon
- Exogenous glucose
- inhibits both cAMP
- synthesis and the
- uptake of other
- sugars, such as
- lactose.
- Components of the
- cascade
- - HPr, the
- phosphotransferase
- (for histidine protein)
- transfers the
- phosphate from
- IIAGlcP to sugar as
- the sugar is transported.
- - IIAGlc protein has two forms
- IIAGlcP activates adenylate
- cyclase to make cAMP.
- IIAGlc inhibits sugar-
- specific permease that
- transport sugar
22
Upstream activator site (UAS)
1. The aCTD (carboxyl terminus of the a
subunits) binds to UP element (UAS),
and aNTD binds to subunit. (A, B)
2. Some promoters lack a -35 sequence and instead
have what is called extended -10 sequence.
This sequence is recognized not bys4 but,
rather by s3. (C)
23
Hypothesis for CAP-cAMP activation of lac
transcription
24
Proposed CAP-cAMP Activation of lac Transcription
- The CAP-cAMP dimer binds to its target site on
the DNA - The aCTD (a-carboxy terminal domain) of
polymerase interacts with a specific site on
CAP(ARI activation region I) - Binding is strengthened between promoter and
polymerase
- (The asubunit N-terminal and C-terminal domains
(a-NTD and aCTD, respectively) fold
independently to form two domains that are
tethered together by a flexible linker.)
25
V. Regulation of gene expression
- 2. Posttranscriptional regulation Gene
expression can be - regulated by
- (1) Inhibition of the translation of the
gene even after mRNA is - made (translational regulation).
- (2) Degradation of mRNA as soon as it is
made or before it can - be translated .
- (3) The protein product may be degraded by
other protein, - called protease.
- (4) By feedback inhibition The final
product inhibits enzyme - activity of the first reaction in a
pathway.
26
V. Regulation of gene expression
- 3. Introns and inteins
- (1) some genes have intervening sequence in
the region of - DNA encoding a RNA or protein. These
sequence can move - from one DNA to another. These
sequences must be - spliced out of RNAs and proteins after
they are made to - restore the function of RNAs or
proteins. - i. The intervening sequences that
splice themselves out of - RNA are called introns which are
much more common in - eukaryotic cells.
- ii. The intervening sequences that
splice themselves out of - protein are called intein.
27
Feedback inhibition regulation
28
VI. Expression vectors
- _at_ The cloning vectors designed to express (made)
large amounts - of proteins for biochemical or structural
analysis. - Besides the elements of cloning vectors,
expression vectors - should have a promoter including operator,
TIR including ATG, - SD sequence and termination codon.
- The gene or DNA sequence inserts into cloning
site must be - in-frame with ATG.
- For easy purification of expressed protein, some
affinity tags - are also include in the vectors.
- (1) Histidine tag DNA sequence encoding
six histidine amino - acids
- i. Histidines binds strongly to
nickel, and so the protein - contains histidines will bind to a
column containing nickel. - ii. Then the bound protein can be
eluted by washing the - column with high concentration of
imidazole, which also - binds to nickel and so will
displace the Hist tag. - (2) Other tag, such as glutathione
S-transferase (GST) is used - often.
29
VI. Expression vectors
- Use pET-15b as an example.
30
VI. Expression vectors
31
Transcriptional and tranlational fusions to
express lacZ
32
VII. Some methods for studying gene
expression - Northern blotting
- Buffer (20 X SSC) /1 L, pH 7.0 175.3 g of
sodium - chloride 88.2 g 0f sodium citrate
33
Northern Blots
- You have cloned a cDNA
- How actively is the corresponding gene expressed
in different tissues? - Find out using a Northern Blot
- Obtain RNA from different tissues
- Run RNA on an denatureing agarose gel (usually
- containing formaldehyde) and blot to membrane
- Hybridize to a labeled cDNA probe
- Northern plot tells abundance of the transcript
- Quantify using densitometer
- Cytoplasmic mRNA isolated from 8 rat tissues
probed with GPDH (glyceraldehyde-3-phosphate
dehydrogenase)
34
VII. Some methods for studying gene
expression Reverse transcription
35
VII. Some methods for studying gene
expression - Primer extension
- Start with in vivo transcription, harvest
cellular RNA containing desired transcript - Hybridize labeled oligonucleotide 18nt (primer)
- Reverse transcriptase extends the primer to the
5-end of transcript - Denature the RNA-DNA hybrid and run the mix on a
high-resolution DNA gel - Can estimate transcript concentration also
36
VII. Some methods for studying gene
expression - S1 nuclease mapping
- Use S1 nuclease mapping to locate the ends of
RNAs - and to determine the amount of a given RNA in
cells at - a given timeLabel a ssDNA probe that can only
- hybridize to transcript of interest
- - Probe must span the sequence start to finish
- - After hybridization, treat with S1 nuclease
which - degrades ssDNA and RNA
- - Transcript protects part of the probe from
- degradation
- - Size of protected area can be measured by gel
- electrophoresis
- Amount of probe protected is proportional to
- concentration of transcript, so S1 mapping can
be - quantitative
37
S1 Mapping the 5 End
38
Real-Time PCR
- Real-time PCR quantifies the
- amplification of the DNA as it occurs
- As DNA strands separate, forward and reverse
primers anneal to DNA strand as that in regular
PCR reaction. - A fluorescent-tagged oligonucleotide binds to
part of one DNA strand
39
Fluorescent Tags in Real-Time PCR
- 1. This fluorescent-tagged
- oligonucleotide serves as a reporter
- probe
- Fluorescent tag at 5-end
- Fluorescence quenching tag at 3-end
- 2. With PCR rounds, the 5 tag is
- separated from the 3 tag
- 3. Fluorescence increases with dNTPs
- incorporation into DNA product
- 4. The whole process takes place
- inside a fluorimeter that measure of
- the fluorescence of tag, which is in
- turn is a measure of the progress
- of the PCR reaction (in real time)
- 5. The reaction can be coupled to RT-
- PCR
40
VII. Some methods for studying gene
expression Biochip (Microarray )
41
Run-Off Transcription
- DNA fragment containing gene to transcribe is cut
with restriction enzyme in middle of
transcription region - Transcribe the truncated fragment in vitro using
labeled nucleotides, as polymerase reaches
truncation it runs off the end - Measure length of run-off transcript compared to
location of restriction site at 3-end of
truncated gene - Size of run-off transcript locates transcription
start site - Amount of transcript reflects efficiency of
transcription
42
Nuclear Run-On Transcription
- Isolate nuclei from cells, allow them to extend
in - vitro the transcripts already started in vivo
in a - technique called run-on transcription
- RNA polymerase that has already initiated
- transcription will run-on or continue to
elongate - same RNA chains
- Effective as initiation of new RNA chains in
isolated - nuclei does not generally occur, one can be
fairly - confident that any transcription observed in
the - isolated nuclei is simply a continuation of
- transcription that was already occurring in
vivo - Therefore, the transcripts should reveal not only
- transcription rates but also give an idea
about which - genes are transcribed in vivo.
43
VII. Some methods for studying gene
expression RNA interference (RNAi)
- 1. Also called cosuppression and
posttrancriptional gene - silencing (PTGS)
- 2. RNA interference occurs when a cell encounters
dsRNA from a - virus, a transposon, or a transgene (or
experimentally added - dsRNA).
- 3. This trigger dsRNA Is degraded into 2123-nt
fragments (siRNA) - by an RNaseIII-like enzyme, Dicer.
- 4. The double-stranded siRNA, with Dicer and the
associated - protein R2D2, constitute a complex (complex
B). - 5. Complex B delivers the siRNA to the RISC
loading complex - (RLC), which probably separates the two
strands of the siRNA - and transfers the guide strand to the
RNA-induced slicing - complex (RISC), which includes a protein
called - Argonaute2 (Ago2).
44
VII. Some methods for studying gene
expression RNA interference (RNAi)
- 6. The guide strand of the siRNA then base-pairs
with - the target mRNA in the active site in the
PIWI - domain of Ago2, which an RNase H-like enzyme
- also known as slicer.
- 7. Slicer cleaves the target mRNA in the middle
of the - region of its base-pairing with siRNA.
- 8. In an ATP-dependent step, the cleaved mRNA is
- ejected from the RISC, which can then accept
a new - molecule of mRNA to be degraded.
45
RNA interference (RNAi)
46
RNA interference (RNAi)
- shRNA (siRNA) hRluc (Renilla luciferase)
Recommended
CrystalGraphics Presentations





























