
Island Biogeography - PowerPoint PPT Presentation
1 / 21
Title:
Island Biogeography
Description:
Determinants of extinction rate ( S ) Should increase with S (more species greater the rate than ... also account for lower S* on more distant islands of ... – PowerPoint PPT presentation
Number of Views:1154
Avg rating:3.0/5.0
Title: Island Biogeography
1
Island Biogeography
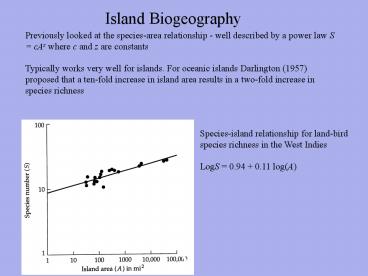
Previously looked at the species-area
relationship - well described by a power law S
cAz where c and z are constants Typically works
very well for islands. For oceanic islands
Darlington (1957) proposed that a ten-fold
increase in island area results in a two-fold
increase in species richness
Species-island relationship for land-bird species
richness in the West Indies LogS 0.94 0.11
log(A)
2
How can we explain higher species richness on
larger islands? Greater habitat
diversity? Large West Indian Islands (Cuba,
Hispaniola, Jamaica) have substantial areas of
montane habitat (mountains over 2000 m) and
consequently windward (wet) and leeward (dry)
slopes Intermediate sized islands are volcanic
plugs that lack habitats of large islands (eg
swamplands, dry limestone forests). Small
islands are coral atolls with simple vegetation
structure. Might work for the Caribbean - but
what about differences in diversity in same size
islands elsewhere?
3
After Darlington, it was recognized that
distance to the mainland also affects species
richness (distance effect)
Diamond (1972) compared species richness on
islands with that expected for an island near
(
Guinea Islands Tropical Pacific
4
Equilibrium theory of Island Biogeography
(Munroe 1948, MacArthur and Wilson 1963,1967).
(See Gotelli (2001) for good description)
Accounts for island size and distance from
mainland Species number on an island represents
the balance between recurrent immigration of new
species onto an island and recurrent extinction
of existing species. Key assumption of the
model There is a permanent mainland source of
species somewhere, P, from which colonists
arrive Immigration rate ?s number of new
species colonizing per unit time Extinction rate
µs number of species going extinct per unit
time dS/dt ?s - µs
5
Determinants of immigration rate
Maximum immigration rate (I) occurs when island
is empty and decreases as more species are added
(so that fewer species remain in the source pool
as potential colonists) Once all the potential
colonists are on the island then S (species
richness on the island) P (mainland source
pool) and immigration rate must 0
Immigration rate ?s Intercept slope(S) I
(-I/P)S
6
Determinants of extinction rate (µS ) Should
increase with S (more species greater the rate
than they can disappearMaximum extinction rate
(E) occurs when all species from the source pool
are on the island (when S P), and must be zero
when no species are present
Intercept 0, therefore extinction rate slope
S µS (E/P)S
7
Substitute in linear terms for immigration and
extinction into formula for rate of change in
species richness dS/dt I-(I/P)S -
(E/P)S Solve for equilibrium species richness
S IP/(IE) determined by size of source pool
and max. immigration and extinction rates...
S is point at which rate of arrival of species
is exactly matched by rate of extinction S has a
characteristic T - the rate of turnover of
species per unit time at equilib.
8
Turnover is a key feature of this model because
there is no fixed stable composition of species -
species composition continues to change although
species number remains relatively constant
Notice on previous graph that T/S E/P T
SE/P Substituting back the term for S T
((IP/IE)E)/P T IE/(IE) or turnover rate
depends only on the maximum immigration and
extinction rates (NOT on the species pool
size) Notice this model doesnt (so far) explain
the species area relationship. What do we need
for that?
9
Two more assumptions to add to the
model Consider two islands of different
size but equal distance from pool
- Larger islands support larger population sizes
of individual species - Probability of species
extinction decreases with increasing population
size
Es max. extinction rate on small island El
rate on large island Immigration curve same for
both islands as both are the same distance away
from the mainland species pool
10
Can also account for lower S on more distant
islands of the same size by changing immigration
rate
In immigration rate for islands close to
species pool rate for far island If Turnover
rate of species lower for far island
No biology in this theory!! Species richness
determined solely by area of the island
(extinction) and distance from mainland
(immigration)... Species have identical traits
11
What are the assumptions of the theory?
Species have similar colonization and
extinction rates Population sizes scale with
island size (no effect of species richness on
population size via competition) Immigration
rate inversely proportional to distance (likely
to be true?) Probability of extinction is
inversely proportional to population size
Probability of immigration and extinction is
independent of species composition on the island
(ie no species interactions). Some assumptions
may not significantly alter the model predictions
(e.g. non linear immigration rates - might be
expected if species differ in their dispersal
characteristics, or might expect extinction rates
to increase with increasing species richness -
why?)
12
Model predictions are fairly robust to non-linear
extinction and immigration functions and were
incorporated into the original model
13
More problematic assumptions
- Isolation does not affect the extinction rate on
an island - Rescue effect
- 2. Size does not affect the immigration rate
- Target effect
14
Rescue effect higher rates of continued
immigration of individuals on near versus far
islands will result in higher population sizes
(or more patches of populations) and potentially
greater genetic diversity. Both factors may
reduce extinction rates (Brown and Kodric-Brown
1977). Lower extinction rate for near islands
will result in lower than expected species
turnover rate
(Extinction rate without rescue effect)
(Lower extinction rate for near island reduces
predicted T)
15
Target effect Island size also likely to
influence immigration rate. Large islands present
larger targets to which immigration can
successfully occur. Results in prediction of
greater turnover rate on large islands.
Immigration rate with target effect
Immigration rate no target effect
16
Target effect raises question of whether one
might expect to see higher species richness on
large islands simply because of higher
colonization (with no need to invoke lower
extinction rates).
This was addressed by Coleman (1982) using the
passive sampling model - which relates
probability of occupancy of a species to the
relative area of a given island in an archipelago
of islands. Species disperse to islands and
accumulate (no extinction) Predicts higher
richness on larger islands, and that abundant
species on the mainland would be common on large
islands. Species that are rare on the mainland
would be rare or absent on small islands
17
How well supported are the assumptions and
predictions of the MacArthur-Wilson theory? 1.
Variation in immigration and extinction
rates Even apart from the consideration whether
extinction rates and immigration rates vary
linearly or non-linearly with species richness,
what evidence is there that they vary at all with
S?
18
Not many measurements of immigration/extinction
rates in published literature Williamson (1981)
analyzed data on bird populations in a plot
within 16 ha oak wood in UK (Eastern Wood)
censused annually 1947-1975. Looked at
extinction/immigration as a function of species
breeding each yr.
Extinction rate tended to increase with species
richness (but not significant)
Immigration rate did decline with increasing
species richness
19
Extinctions and immigrations are not
equiprobable. Core group of 14 spp breed in the
wood every year. Extinctions and immigrations
occur among a group of 11 species with transient
populations Williamson (1981) Plant colonization
to Surtsey (volcanic island 33km south of iceland
- appeared in 1963). Immigration highest after
initial pioneers established?
Species present
20
2. Variation in species population size with
island size Seems intuitively reasonablebut
good reason to think that island size will not be
the primary determinant of population size in
many cases - ecological release from predators
or competitors may allow some taxa to maintain
higher population densities on small vs large
islands/mainland when interacting species are
absent. Crowell (1983) introduced deer mice and
onto rodent-free islands in the Deer Island
Archipelago, Maine. Populations grew to greatly
exceed mainland densities Case (1975) Lizard
densities on islands in Gulf of California
highest on small islands. Lizard density actually
declined with increasing island area and island
species richness.
21
MWs equilibrium model is distinguished from
passive sampling colonization models and
community assembly based on species interaction
by virtue of predicting substantial species
turnover. Turnover however is difficult to
assess (Friday discussion). High presumed rates
of species turnover tend to be largely the result
of cycling of transient peripheral species. If
transient species are excluded then turnover
often very low. Departures from predictions of
the MW island biogeography null model may be
the most important contribution of this theory
Looking at how islands work compared to the
mainland provides an important investigative tool
in community ecology.
3. Substantial species turnover at equilibrium
Recommended
CrystalGraphics Presentations





























