
Amino Acids and Proteins PowerPoint PPT Presentation
1 / 37
Title: Amino Acids and Proteins
1
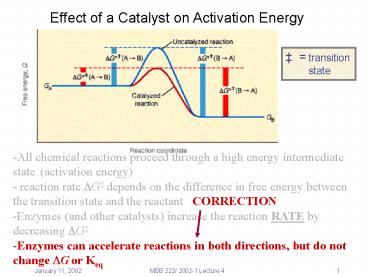
Effect of a Catalyst on Activation Energy
transition state
-All chemical reactions proceed through a high
energy intermediate state (activation energy) -
reaction rate ?G depends on the difference in
free energy between the transition state and the
reactant CORRECTION -Enzymes (and other
catalysts) increase the reaction RATE by
decreasing ?G -Enzymes can accelerate reactions
in both directions, but do not change ?G or Keq
2
Proteins I Amino Acids Polypeptides Folding
1. Amino Acids and their Chemical
Properties 2. Peptides Levels of Protein
Structure i) Secondary Structure ?-helices,
?-sheets and ?-turns ii) Tertiary and Quaternary
Structure Coiled-Coils etc. iii) Protein
folding iv) Methods to Determine Protein
Structure X-ray crystallography v) Simple
Computer Visualization of Protein Structure
3
Figure 5.1 3D Structure of Myoglobin
- first to be determined by x-ray crystallography
- revealed how the protein bound heme (loaded
with oxygen) and gave the first detailed look at
a protein structure
- now over 2000 protein structures are known
4
Amino Acid structure
Side chain
? - carbon
Carboxyl group
Amino group
While most free amino acids have a net zero
charge at neutral pH, they have both and
charges. Zwitterion
NH3
COO-
5
Acidic and Basic Amino Acids- Hydrophilicmost
often found at the outer surface of proteins,
H-bonding
- these are charged at neutral pH (physiological
conditions)- contribute to charge of protein
6
These have uncharged amide and hydroxyl groups
that participate in H bonding
7
Hydrophobic amino acids
Insoluble in water, cluster on the insides of
proteins
Nonpolar, hydrocarbons
Aromatic side chains
8
(No Transcript)
9
Proline
Proline is a special amino acid in fact, it is
more appropriately named an imino acid - it is
the only group that is cyclic, where the side
chain forms a covalent bond with the nitrogen atom
10
Glycine (Gly) G
H H2NCCOO- H
- smallest and most versatile of amino
acids can readily fit into any protein
structural element
11
Cysteine side chains can form a disulfide bond to
form cystine often plays important role in
protein structure
12
Amino acid structures
aspartic acid (D)
glutamic acid (E)
valine (V)
leucine (L)
methionine (M)
isoleucine (I)
phenylalanine (F)
tyrosine (Y)
tryptophan (W)
aspargine (N)
glutamine (Q)
serine (S)
threonine (T)
glycine (G)
lysine (K)
histidine (H)
arginine (R)
cysteine
proline
alanine (A)
2
3
know all of them for this course!
13
Amino acids in proteins
- Average molecular weight is 113- allows you to
calculate how many amino acid residues are in a
protein of a given size - Leucine, serine, lysine and glutamic acid are the
most abundant- constitute 32 of amino acids in
proteins - Cysteine, tryptophan, methionine are rare-
together total 5
14
Stereoisomers of Amino Acids
All of the Amino Acids incorporated by organisms
into proteins are in the L form
Note glycine is not chiral because of H- being
its side chain.
15
Amino acids are weak acids bases
-all free amino acids are acids and bases because
of free carboxyl and amino ends - in peptides
and proteins these are involved in peptide bonds
so in proteins only some of the amino acids
posses acid/base properties - in the
three-dimensional environment of proteins the
pKas of the amino acids can differ significantly
from the values in table 5.1 (for the free amino
acids)
Table 5.3
16
Peptide bond formation
The peptide bond has partial double bond character
-
O
C
N
H
Resonant Structures
17
Planar Character of a Peptide Bond
Figure 5.12 - partial double bond character
prevents the peptide bond from rotating
18
Figure 6.2 rotation about the alpha carbon
- with the lack of rotation around the peptide
bond the point of flexibility along the backbone
of the protein arises at the ? - carbon.
?-carbon
Bonds allowing for rotation along the protein
backbone
19
Figure 6.27 - Levels of Protein Structure
The folded protein structure is stabilized by a
variety of weak chemical interaction, and in
some cases covalent (disulfide) bonds between
cysteine residues
- - helix
- Myoglobin
- Hemoglobin
Disulfide bond
R CH2SSCH2R
Cys Cys
20
Protein structure overview
Structural element Description primary
structure amino acid sequence of protein
secondary structure helices, sheets,
turns/loops super-secondary structure associatio
n of secondary structures domain self-containe
d structural unit tertiary structure folded
structure of whole protein includes
disulfide bonds quaternary structure assembled
complex (oligomer) homo-oligomeric (1
protein type) hetero-oligomeric (gt1
type)
21
Protein secondary structure helices
- rod like right-handed
INTRA-chain H-bonds between gtCO group of each
peptide residue and the gtN-H group of the 4th
amino acid away
- alpha helices are about 10 residues on average
H-bonding
- side chains of alpha-helices are well
staggered, preventing steric hindrance
- helices can form bundles, coiled coils, etc.
22
Helical Wheels
- a tool to visualize the position of amino acids
around an alpha-helix - allows for quick
visualization of whether a side of a helix posses
specific chemical properties - example shown is
a helix that forms a Leucine-Zipper
23
?-Helix Breakers
Most amino acids like to be in an
?-helix. Notable exceptions GLYCINE PROLINE
(Imino Acid)
No Hydrogen On this N to H-Bond
O
C-O
N
H
- proline residues often serve as ?-Helix
Breakers - often found at the boundaries of
?-Helices and in turns
24
Proteins with ?-helices
Major structural component in many proteins, some
globular proteins contain mostly ?-helices,
connected by turns (i.e., hemoglobin 70
?-helices)
Some Interesting ?-Helices - small DNA binding
helices - membrane spanning helices -
amphipathic helices - coiled Coils
25
DNA Binding - an ??-helix fits perfectly into the
major groove of double stranded DNA. - many DNA
binding proteins use particular ??-helices to
specifically recognize a DNA sequence.
dsDNA
Membrane Spanning - contains hydrophobic amino
acids in the central region to allow the protein
to cross a bi-layer membrane
Hydrophilic Hydrophobic
26
Amphipathic Helices Amphipathic hydrophilic
hydrophobic - these helices posses hydrophilic
amino acids on one side and hydrophobic residues
on the other. - these ?-helices in some cases
can be used to associate a protein to a membrane.
Hydrophobic
Hydrophilic
hydrophilic head group
aliphatic carbon chain
lipid bilayer
27
6.13 Coiled Coils a higher order structure
composed of alpha-helices
Alpha-Helix
A Triple Coiled Coil
Axis for Interaction with other alpha-helix. In a
double coiled-coil.
28
Alpha-Helix Beta-Sheet
29
Protein structure beta-sheets
- the basic unit of a beta-sheet is called
a beta-strand
- repeating unit like the alpha helix
- beta-sheets can form various higher-level
structures, such as a beta-barrel
parallel
twisted
anti-parallel
30
The Beta-Sheets
- - strands of amino acids held together in sheets
by INTER-STRAND H-Bonding - - bonding between backbone gtCO and gtN-H on
different strands - - sheets tend to be curved
- - the R-groups lie perpendicular to the sheets
stick out on either face of the sheet
R
R
R
R
R
R
R
R
-variable polarity etc
R
R
R
31
Beta-Sheets and DNA
- an alpha-helix is of appropriate size to fit in
the major groove of DNA - beta sheets fit very
well into the minor groove of DNA double
helices - beta-sheets can also used in DNA
binding but are generally less commonly used
Alpha-Helix
Beta-Sheet
32
Figure 6.12 An example of complex
beta-sheets Silk Fibroin - multiple pleated
sheets provide toughness rigidity to many
structural proteins.
33
Protein structure turns/loops
beta-sheet
alpha-helix
- there are various types of turns, differing in
the number of residues and H-bonding pattern
- loops are typically longer they are often
called coils and do not have a regular, or
repeating, structure
loop (usually exposed on the surface of proteins)
34
Beta - Turns
Figure 6.18
There are two classes of beta-turns - type I -
type II Note the position of R2 and R3 in both
cases Type I turns have the amino acids on the
same side. Type II turns have the amino acids on
the opposite sides Note H-bonding between
backbones of residue 1 4
35
Gamma-Turns
Proline
A 3 amino acid turn utilizing proline at the
turn. H-bonding with CO of residue 1 and N-H of
residue 2
36
Transitions between Secondary Structure Elements
- at the end of a domain or secondary structure
the amino acids can turn in various ways - some
turns are not allowed - this is due the steric
interference. - some other types of turns a
quite common because of their stability, because
H-bonding in their structure
Figure 6.9
37
Folding of RNAse A in the test tube
denaturation
renaturation
Incubate protein in guanidine hydrochloride (GuHCl
) or urea
100-fold dilution of protein into
physiological buffer
- the amino acid sequence of a polypeptide is
sufficient to specify its three-dimensional
conformation Thus protein folding is a
spontaneous process that does not require the
assistance of extraneous factors
Recommended
