
Diapositiva 1 - PowerPoint PPT Presentation
1 / 30
Title:
Diapositiva 1
Description:
... exhiben las caracter sticas esenciales de las c lulas madres embrionales (CME). Estas c lulas madres pluripotentes humanas inducidas tienen cariotipos normales, ... – PowerPoint PPT presentation
Number of Views:82
Avg rating:3.0/5.0
Title: Diapositiva 1
1
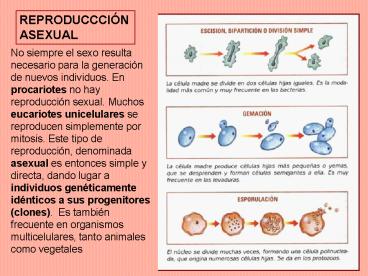
REPRODUCCCIÓN ASEXUAL
No siempre el sexo resulta necesario para la
generación de nuevos individuos. En procariotes
no hay reproducción sexual. Muchos eucariotes
unicelulares se reproducen simplemente por
mitosis. Este tipo de reproducción, denominada
asexual es entonces simple y directa, dando lugar
a individuos genéticamente idénticos a sus
progenitores (clones). Es también frecuente en
organismos multicelulares, tanto animales como
vegetales
2
REPRODUCCIÓN SEXUAL
El ciclo reproductivo sexual involucra una
alternancia de generaciones haploides de células
(con una dotación simple de cromosomas) con
generaciones diploides de células, que contienen
un doble juego de cada uno de los cromosomas (los
cromosomas homólogos provistos por cada uno de
los progenitores). La mezcla de genomas se
realiza mediante la fusión de dos células
haploides (las gametas, que contienen un solo
juego de cromosomas) para formar una célula
diploide.
3
REPRODUCCIÓN SEXUAL
Las células haploides especializadas en la fusión
sexual se denominan gametas. La gameta femenina
es usualmente grande e inmóvil en tanto que la
gameta masculina es habitualmente pequeña y
móvil. En general hay dos líneas celulares una
es la línea germinal, de la cual provendrá la
próxima generación de gametas, y la otra está
representada por las células somáticas, que
formarán el resto del cuerpo y que no están
capacitadas para generar descendencia.
Cuáles son los beneficios que aporta? Parece ser
que el reordenamiento genético ayuda a una
especie a sobrevivir ante variaciones ambientales
impredecibles. Si los padres producen muchos
descendientes con una amplia variación de
combinaciones genéticas, hay una mejor chance de
que al menos uno de los descendientes disponga de
la dotación genética necesaria para la
supervivencia en condiciones en que la mayoría no
pueda hacerlo.
4
MEIOSIS
La meiosis implica dos divisiones nucleares
5
División I de la Meiosis
6
División II de la Meiosis
7
Reconocimiento de los cromosomas homólogos
Un núcleo diploide contiene dos versiones muy
similares de cada cromosoma (paterno y materno)
los cromosomas homólogos (a la derecha,
cariograma humano con los homólogos agrupados).
Cuando cada cromosoma es duplicado por
replicación del ADN (fase S) las copias de cada
doble hélice se denominan cromátidas hermanas. En
la mitosis, cada una de las cromátidas hermanas
se separa en la anafase y se convierte en un
cromosoma individual.
La meiosis requiere que cada par de homólogos se
reconozcan y se apareen físicamente antes de que
se ubiquen en el huso mitótico. El reconocimiento
de ambos homólogos y su íntimo contacto hasta la
metafase en que se separan es esencial para
asegurar la distribución equitativa de los
cromosomas homólogos. De no ser así los
cromosomas se distribuirían al azar y una célula
hija podría recibir un par de homólogos y carecer
en absoluto de una copia de otros cromosomas,
distribución que sería letal para ellas
8
(No Transcript)
9
División I de la Meiosis
La nueva progenie producida la división I de la
meiosis contiene por lo tanto una cantidad
haploide de cromosomas, difiriendo de las células
diploides comunes en dos aspectos 1) cada
cromosoma contiene dos copias de ADN, que
provienen de sólo uno de los dos cromosomas
homólogos (a lo que se agrega la posibilidad del
recambio genético) y 2) las dos copias han sido
heredadas como cromátidas hermanas estrechamente
asociadas, como si fueran un único cromosoma
10
Tras una interfase muy corta (intercinesis) tiene
lugar la formación de las gametas, que ocurre a
través de una segunda división, la división II de
la meiosis, sin que se produzca una nueva
replicación del ADN. Las cromátidas hijas se
separan, como en una mitosis normal, para
producir células con un contenido haploide de
ADN. La meiosis consiste así en dos divisiones
celulares con un simple paso de replicación del
ADN, como resultado de lo cual se generan cuatro
células haploides por cada célula diploide que
entra en meiosis.
11
Redistribución génica
A menos que se trate de gemelos, que se
desarrollan a partir de una misma cigota, ninguno
de los hijos de una pareja de progenitores es
igual a otro. Esto se debe a que, antes de que se
produzca la fusión de las gametas, se han
producido dos clases de recomposición genética
durante la meiosis el reordenamiento del genoma
y el entrecruzamiento entre homólogos.
Lisa, Maggie Bart Simpson
Reordenamiento del genoma Es consecuencia de la
distribución al azar de los cromosomas homólogos
maternos y paternos entre las células hijas
durante la anafase de la división I de la
meiosis, como resultado de la cual cada célula
obtiene una mezcla diferente de cromosomas
maternos y paternos. Simplemente por este proceso
de separación al azar de los homólogos uno puede
obtener 2n distribuciones distintas, siendo n el
número haploide de cromosomas así, para la
especie humana cada individuo podrá producir 223
8,4 x 106 gametas diferentes
12
Entrecruzamiento entre Homólogos (Crossing Over)
Ocurre durante la larga profase de división I de
la meiosis, en la que se intercambian partes
equivalentes de los cromosomas homólogos. En
promedio, ocurren dos a tres entrecruzamientos en
cada par de cromosomas. El proceso de
entrecruzamiento implica la ruptura del ADN de
una cromátida paterna y lo propio en la zona
equivalente de una cromátida materna, con el
consiguiente intercambio de fragmentos de una
cromátida a otra
13
Entrecruzamiento entre Homólogos (Crossing Over)
14
Mantiene a los cromosomas homólogos en contacto
lineal y consiste en un núcleo proteico (el
elemento central) en forma de escalera de mano, a
cuyos costados se alinean los dos homólogos. Las
cromátidas hermanas están mantenidas muy juntas a
uno de los dos lados del eje proteico y su ADN se
encuentra parcialmente extendido formando una
serie de bucles. Los nódulos de recombinación
funcionan como máquinas de recombinación
enzimática que acerca regiones equivalentes de
ADN materno y paterno para que el
entrecruzamiento tenga lugar a través del
complejo sinaptonémico
Complejo sinaptonémico
15
Rol de los quiasmas en la segregación de los
homólogos
Los quiasmas son esenciales para asegurar la
correcta segregación de los dos homólogos hacia
los núcleos hijos (papel similar al del
centrómero en la mitosis), manteniendo a los
homólogos paterno y materno juntos en el huso
hasta la anafase I.
Los homólogos con quiasmas se mantienen juntos
debido a que las cromátidas hermanas están
pegadas todo a lo largo por las proteínas
denominadas cohesinas. Estas son parcialmente
digeridas por la separasa al iniciarse la anafase
I pero permanecen al menos a nivel de los
centrómeros.
16
División II de la Meiosis
Es similar a una división mitótica. Luego de la
larga profase I se suceden dos divisiones
celulares sucesivas, sin que exista un período
intermedio de síntesis de ADN. Después de
finalizar la primera división meiótica se vuelven
a formar las membranas alrededor de los núcleos
hijos y comienza una breve interfase. Dado que
no ocurre replicación del ADN, en algunos
organismos los cromosomas parecen pasar casi
directamente de una fase a la otra. En todos los
organismos la profase II es breve la envoltura
nuclear se rompe y se forma un nuevo huso, luego
de lo cual siguen las etapas correspondientes a
la metafase II, la anafase II y la telofase II.
17
Diferencias entre Anafase I y II
En la metafase I de la meiosis los cinetocoros de
ambas cromátidas hermanas parecen haberse
fusionado, de tal modo que las fibras del
cinetocoro tiran todas hacia la misma dirección y
los cromosomas homólogos son mantenidos juntos
únicamente en la zona de los quiasmas .Durante la
división meiótica II, las cromátidas hermanas
están unidas solamente a nivel de los centrómeros
y a ambos lados se unen microtúbulos del
cinetocoro que tiran de las cromátidas en
direcciones opuestas durante la anafase II.
18
No Disyunción meiótica
A veces el proceso de división no se desarrolla
correctamente y puede llevar a células con un
número de cromosomas anormal, pudiendo haber
cromosomas por exceso (trisomía) o por defecto
(monosomía) respecto al número constante de la
especie, con un desequilibrio del material
genético. La no-disyunción en la primera división
meiótica implica la no segregación de un par de
homólogos en la segunda división meiótica
conlleva la no separación de las cromátidas
hermanas.
19
GAMETOGÉNESIS
La meiosis es equivalente tanto en plantas como
en animales, pero la producción de gametas
involucra mecanismos adicionales, que suelen
diferir entre los distintos organismos e incluso
entre ambas gametas.
En los embriones de vertebrados las células
germinales primordiales (precursoras de las
gametas), formarán los ovarios en las hembras y
los testículos en los machos, responsables de la
formación de las gametas (óvulos y
espermatozoides). La fusión del óvulo y el
espermatozoide generará la cigota que dará lugar
al embrión, donde nuevas células se diferenciarán
en germinales primordiales y reiniciarán el
ciclo. Una gónada (órgano reproductor) se
convertirá en ovario a menos que sus contenga un
cromosoma Y, en cuyo caso se convertirá en
testículo. En vertebrado superiores el gen SRY
del cromosoma Y es el responsable de la
determinación de la formación de testículo se
expresa en ciertas células somáticas de la gónada
en desarrollo desencadenando una cascada de
reacciones que generan los espermatozoides y, a
su vez, promueven el desarrollo de las
características masculinas, suprimiendo el de las
femeninas.
20
Ovogénesis
Las células germinales primordiales migran hacia
las gónadas y se transforman en ovogonias, que se
dividen por mitosis, antes de diferenciarse en
ovocitos primarios. Sobreviene luego la primera
división meiótica, en cuya profase el ovocito
primario puede mantenerse de unos días a varios
años antes de proseguir con la división. Durante
esta etapa se sintetiza la cubierta y se acumulan
granos corticales y demás material necesario para
el desarrollo embrionario. La maduración del
ovocito usualmente no ocurre hasta la madurez
sexual, estimulada por hormonas. Bajo este
estímulo genera dos células conteniendo la mitad
del número de cromosomas una célula pequeña
(primer cuerpo polar) y el ovocito secundario. La
segunda división meiótica produce el segundo
cuerpo polar (pequeño y destinado a desaparecer
como el anterior ) y el óvulo maduro. En la
mayoría de los vertebrados la maduración del
ovocito secundario se detiene en la la metafase
II y es liberado del ovario si se produce la
fecundación, es estimulado a completar la
meiosis.
21
Espermatogénesis
La espermatogénesis difiere de la ovogénesis en
que no comienza hasta la pubertad (depende de la
secreción de la testosterona) y se produce de
manera continua en la capa epitelial de los
túbulos seminíferos, dentro de los testículos.
En la capa más externa del túbulo seminífero se
encuentran las espermatogonias, que luego se
diferencian en espermatocitos primarios. Estas
células sufren la primera división meiótica para
dar dos espermatocitos secundarios, haploides y
con un cromosoma X o un Y. Los dos
espermatocitos primarios sufren la segunda
división meiótica y generan cuatro espermátidas,
con un número haploide de cromosomas. Estas
espermátidas todavía deben diferenciarse
(espermiogénesis) en espermatozoides, que son
conducidos hacia el epidídimo, un tubo que rodea
los testículos, donde sufren la maduración final
y son almacenados.
22
Espermatogénesis
23
Fecundación
24
Fecundación
25
Fecundación
26
Embriogénesis
Luego de la fecundación del óvulo, la cigota
experimenta una serie de divisiones rápidas, en
las cuales sólo se duplica el ADN. A partir del
estadio de 16 células el embrión adquiere la
forma de una esfera sólida (mórula) y luego se
convierte en una esfera hueca (blástula), en la
que se diferencian el macizo celular interno,
primordio del futuro cuerpo y el trofoblasto que
interviene en la formación de la placenta.
Posteriormente el macizo celular interno da lugar
a un embrión discoide plano con tres capas
epiteliales superpuestas el ectodermo, el
mesodermo y el endodermo.
27
Embriogénesis
Dado que las células diferenciadas más primitivas
no escapan a la regla de poseer los mismos genes,
las desigualdades iniciales entre ellas deben
buscarse en el citoplasma que heredan de la
célula huevo. En efecto, se considera que el
citoplasma de la cigota contiene moléculas
distribuidas asimétricamente, que llevan el
nombre de determinantes citoplasmáticos del
desarrollo, los cuales se reparten de manera
desigual entre las células del embrión. Se cree
que estos determinantes son proteínas que actúan
como factores de transcripción específicos
28
(No Transcript)
29
Líneas de Células Madre pluripotentes inducidas
derivadas de células somáticas humanas
Junying Yu y colaboradores, Science, vol. 318,
págs. 1917-20, 21 de diciembre de 2007
La transferencia al núcleo de una célula somática
de factores presentes en el oocito de mamíferos
permite reprogramar el núcleo de la célula
somática a un estado indiferenciado. Hemos
demostrado que cuatro factores (OCT4, SOX2,
NANOG, and LIN28) son suficientes como para
reprogramar células somáticas humanas a células
madres pluripotentes que exhiben las
características esenciales de las células madres
embrionales (CME). Estas células madres
pluripotentes humanas inducidas tienen cariotipos
normales, expresan actividad telomerasa, poseen
marcadores celulares de superficie y genes que
caracterizan a las CME humanas, manteniendo el
potencial de desarrollo para diferenciarse en
células derivadas avanzadas de las tres capas
germinales primarias. Tales líneas celulares
humanas pluripotentes serían útiles en la
producción de nuevos modelos de enfermedades y en
el desarrollo de fármacos, así como en
aplicaciones para medicina de transplantes, luego
que algunas limitaciones técnicas (por ej. las
mutaciones a través de la integración viral) sean
eliminadas.
30
Clonación terapéutica vs. Reprogramación del
Núcleo
La clonación terapéutica crea células madre que
pueden desarrollar distintos tejidos del
organismo humano. Esta técnica supone la creación
y destrucción de embriones, lo que suscita una
fuerte controversia ética. Además, las células
madre pueden ser rechazadas por el cuerpo. El
nuevo método evita ambos obstáculos, y es más
barato.
Recommended
CrystalGraphics Presentations





























